编译:微科盟-赵大壮,编辑:微科盟悄咪咪、江舜尧。
导读
动物性食品在消费者日常饮食中所占的比例不断增加,鸡肉因其高蛋白、低脂肪的特点而备受青睐。世界各地的鸡肉消费量正在稳步增长。肌内脂肪(Intramuscularfat,IMF)是影响肉品质的一个关键指标。
目的:提高IMF含量有助于提高鸡肉品质。鸡肉中IMF沉积的调控机制尚不清楚,对其进行全面的阐明对改善鸡肉品质至关重要。
方法:对516只黄羽鸡进行全基因组重测序,对3只63日龄雌性JXY鸡进行单细胞RNA测序。此外,还对不同发育阶段的JXY鸡的胸肌组织进行了转录组测序。应用13C同位素示踪技术。
结果:本研究通过对一个IMF选择群体和一个对照群体的大规模遗传分析,确定脂肪酸合成酶(fattyacidsynase,FASN)是提高IMF含量的关键基因。此外,与传统观点相反,从头脂肪生成(denovolipogenesis,DNL)被认为是IMF沉积的重要贡献者。
论文ID
原名:Genomicinsightsintothecontributionofdenovolipogenesistointramuscularfatdepositioninchicken
译名:鸡肌内脂肪沉积中脂肪新生作用的基因组研究
期刊:JournalofAdvancedResearch
IF:10.7
发表时间:2023.12
DOI:10.1016/
实验设计
结果
1脂肪酸代谢相关基因参与IMF沉积
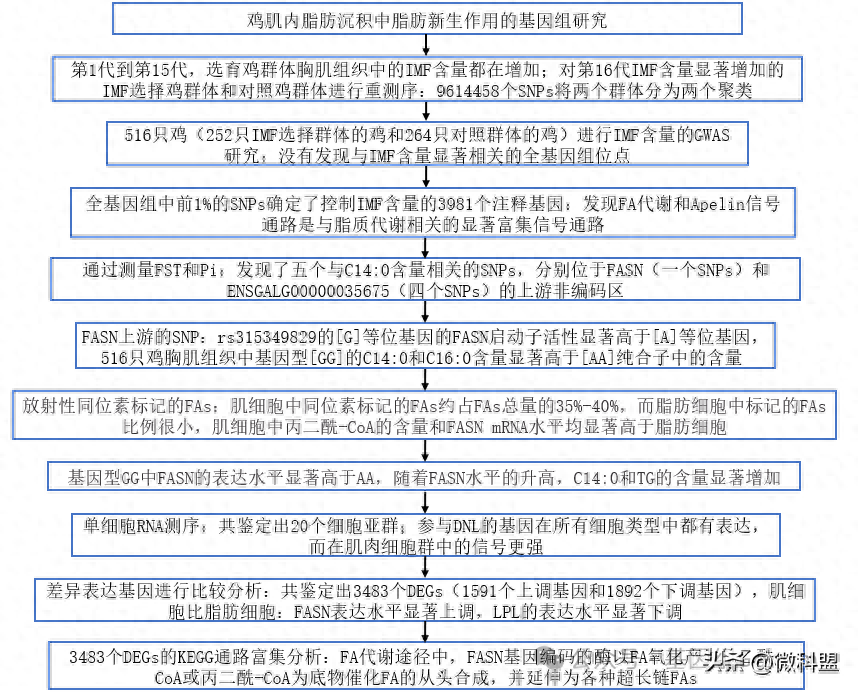
研究人员发现,从第1代到第15代,选育鸡群体胸肌组织中的IMF含量都在增加(图1a)。然后,研究人员对第16代IMF含量显著增加的IMF选择鸡群体和对照鸡群体进行了重测序。基于质量控制后总共9614458个单核苷酸多态性(singlenucleotidepolymorphisms,SNPs),通过群体结构分析将IMF选择的群体和对照群体分成两个聚类(图1b)。此外,在多条染色体(包含845个注释基因)上发现了根据前1%阈值用固定指数(fixationindex,FST)检测到的显著选择信号(图1c)。这表明表型选择导致了基因组分化。此外,经过通路富集分析和筛选,确定了一些显著富集的脂质代谢相关通路,包括甘油脂类代谢、甘油磷脂代谢、PPAR和脂肪细胞因子信号通路。通过对表型数据的统计分析研究了IMF沉积的遗传基础后,研究人员选择了516只鸡(包括252只来自IMF选择群体的鸡和264只来自对照群体的鸡)进行IMF含量的GWAS研究。然而,在全基因组显著性阈值(-log10p=8.28)或提示性阈值(-log10p=6.98)下,研究人员没有发现与IMF含量显著相关的全基因组位点(图1d)。相反,研究人员利用全基因组中前1%的SNPs(n=98869个)确定了控制IMF含量的候选基因。根据所发现的3981个注释基因,发现脂肪酸(FattyAcids,FA)代谢和Apelin信号通路是与脂质代谢相关的显著富集信号通路(图1e中突出显示)。其中,FA代谢通路主要参与FA的生物合成(由ACLY、ME、ACACA和FASN等编码的酶催化)、碳链延长(包括ELOVL5编码的脂肪酸延长酶5等)、酰化(包括ACSL3、ACSL4等编码的酶)和氧化(包括ACADL等编码的酶)。
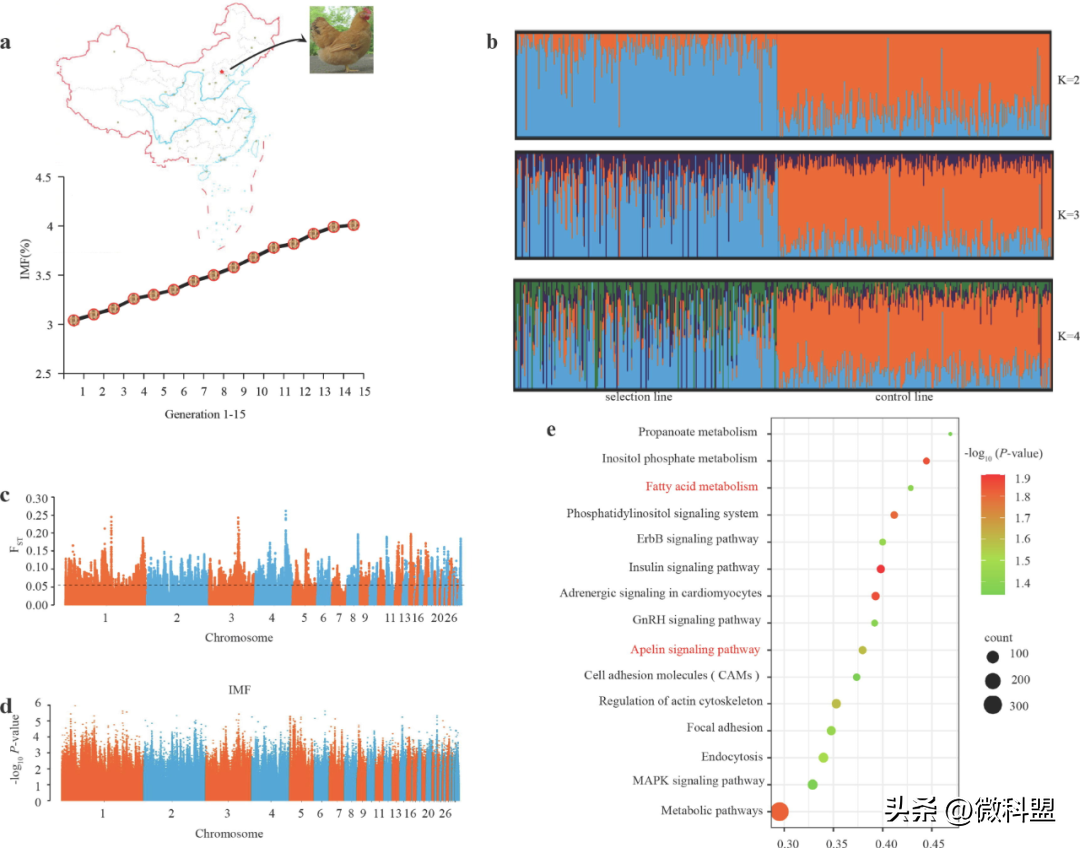
图1|全基因组鉴定与IMF沉积相关的基因。选择品系1~15代的胸肌组织中IMF的含量呈持续增加趋势。选择群体和对照群体(K=2、3、4)的种群结构分析,显示两个种群的分离。c.根据最高1%的阈值,固定指数(fixationindex,FST)显示出显著的选择标记,遗传分化明显(n=516)。黑色虚线代表最高1%阈值线。d.根据全基因组显著性阈值(log10p=8.28)或提示性阈值(log10p=6.98)对IMF含量进行GWAS分析,没有发现与IMF含量相关的全基因组位点(n=516)。e.基于前1%SNPs的3981个注释基因的富集信号通路(n=98869)。
2选择与胸肌肉豆蔻酸和软脂酸含量相关的FASN和rs315349829
考虑到上述结果以及FAs对IMF沉积的重要贡献,研究人员对FAs组成进行了GWAS分析,发现18号染色体上的一个位点与胸肌组织中肉豆蔻酸(C14:0,FAs从头合成的中间产物)的含量显著相关(图2a和2b),该位点还与棕榈酸(C16:0,FAs从头合成的最终产物)的含量相关(图2b)。为避免遗漏有用信息,研究人员将该区域扩大到1.251Mb(chr18:4548962-5800023),阈值线为±20kb,其中包含14938个SNPs,它们具有更高更强的连锁关系。针对选择群体中肉豆蔻酸和棕榈酸含量明显高于对照群体的情况,研究人员用10kb窗口的FST进行了选择性扫描,但只发现了一个弱选择区域(图2c)。研究人员通过测量FST(标准值为0.1)和Pi(图2d)对该区域单个位点的遗传选择信号进行了调查,发现了五个与C14:0含量相关的SNPs,分别位于FASN(一个SNPs)和ENSGALG00000035675(四个SNPs)的上游非编码区(图2e)。根据研究人员报道的JXY鸡转录组数据,FASN基因在胸肌组织中表达,而不在ENSGALG00000035675中表达(图2f)。因此,发现FASN上游的SNP(chr18:4910969,rs315349829[A/G])与重要区域的最高SNP(chr18:4910989,rs312544499[G/A])紧密相连,距离为20bp。通过分别转染携带相应基因的载体进行双荧光素酶报告基因检测发现,rs315349829的[G]等位基因的FASN启动子活性显著高于[A]等位基因(图2g)。研究人员还发现,携带rs315349829的交替(alt)-基因型[GG]的516只鸡胸肌组织中C14:0和C16:0的含量显著高于携带参考(ref)-基因型[AA]的纯合子中的含量(图2h),这表明rs315349829对JXY鸡胸肌组织中C14:0和C16:0的生物合成有促进作用。此外,研究人员还研究了rs315349829对其他FAs的影响,发现除C14:0、C16:0外,alt基因型【GG】纯合子中C14:1、C16:1、C20:1和C21:0的含量显著高于ref基因型【AA】。此外,估计的表型变异表明,在JXY鸡胸肌组织的全基因组遗传变异中,SNPrs315349829对肉豆蔻酸含量的贡献率为27.74%。
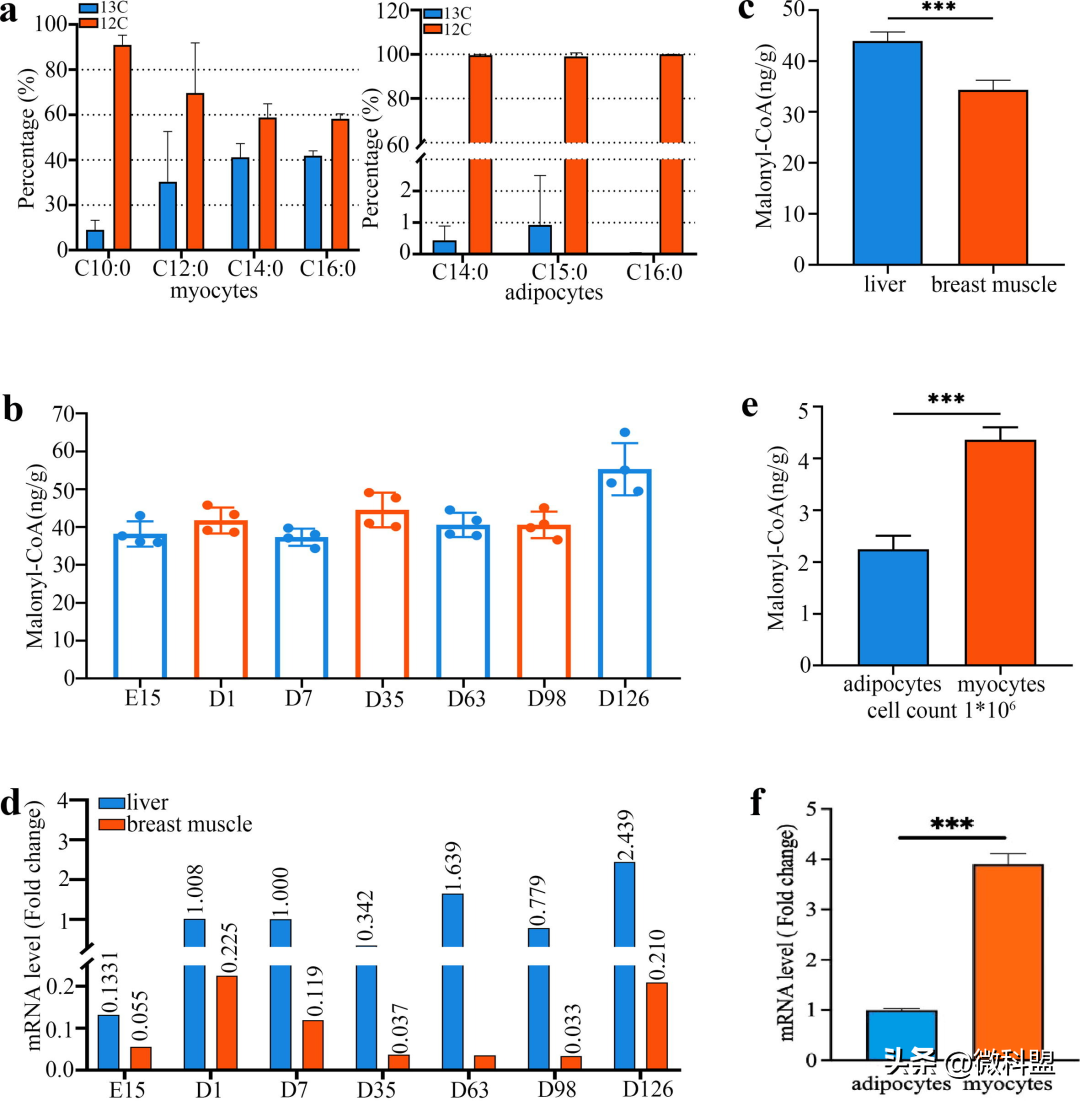
图2|鉴定与胸肌中脂肪酸含量有关的基因变异。鸡肉中豆蔻酸含量的曼哈顿图。红线和蓝线分别代表基因组阈值和提示性阈值。b.将区域扩展到1.251Mb(chr18:4548962-5800023)后,18号染色体上的提示性阈值线为±20kb。在18号染色体chr18:4548962-5800023处的10kb窗口进行选择性扫描,只发现了一个弱选择区域。(标准值为0.1)和Pi对18号染色体chr18:4548962-5800023单个位点的选择信号。e.通过五个与C14:0含量相关的SNPs注释的基因。和ENSGALG00000035675在胸肌组织中的表达水平。的不同纯和基因型([G]和[A]等位基因)对FASN启动子表达的调控(n=3个细胞重复)。h.携带SNPrs315349829的[GG]或[AA]等位基因的JXY纯合子的胸肌组织中C14:0和C16:0的含量(n=516)。鸡的IMF选择群体和对照群体中SNPrs315349829不同基因型数量的分布(n=516)。j.选育种群(n=252)和对照种群(n=264)中携带SNPrs315349829的[GG]或[AA]的JXY纯合子的胸肌组织中C14:0和C16:0的含量。*,**,***。
IMF选育群体中C14:0和C16:0含量较高的GG和AG个体数量高于对照JXY鸡群体,而C14:0和C16:0含量较低的AA纯合子个体数量低于对照鸡群体(图2i和2j),这可能是IMF选育群体中C14:0和C16:0含量较高的原因。此外,对rs315349829的分布分析表明,该突变作为一种[A]到[G]的突变广泛存在于世界各地的鸡品种中。因此,研究人员还研究了rs315349829与其他鸡品种,即文昌鸡(Wenchangchicken,WC)、清远麻鸡(Qingyuanmachicken,QYM)和金陵黄鸡E系(ElineofJinlinghuangchicken,E-JLH)的516只鸡的胸肌组织中C14:0含量的关系,发现C14:0含量(r=0.687,P=1.55E-06)和C16:0(r=0.687,P=1.55E-06)显著相关,这支持了rs315349829对多个品种肌肉组织中C14:0和C16:0含量调控的广泛突变效应。
3DNL主要发生在肌肉组织的肌细胞中
在确定了FASN对DNL的影响后,研究人员通过使用放射性同位素标记的FAs技术研究了FAs的代谢和分布,从而进一步研究了DNL是否发生在鸡的肌肉组织中。不出所料,以放射性同位素标记的葡萄糖为起始底物培养48小时后,在肌细胞和脂肪细胞中都发现了同位素标记的FAs(图3a)。此外,研究人员还发现肌细胞中同位素标记的FAs约占FAs总量的35%-40%,而脂肪细胞中标记的FAs比例很小。此外,研究人员还测定了丙二酰-CoA(DNL的不可逆第一产物)的含量。首先,在所有不同发育阶段的肌肉组织中都发现了丙二酰-CoA(图3b),表明鸡肌肉组织中普遍存在DNL。令研究人员惊讶的是,进一步比较发现,肌肉组织中丙二酰-CoA的含量约为肝脏中丙二酰-CoA含量的80%(图3c),在FASNmRNA水平上也观察到类似的模式(图3d)。此外,研究人员还体外测定了肌肉组织中肌细胞和脂肪细胞中丙二酰-CoA的含量和FASNmRNA水平,结果令人惊讶地发现,肌细胞中丙二酰-CoA的含量和FASNmRNA水平均显著高于脂肪细胞()(图3e)。

图3|鸡胸肌组织中存在DNL。a.用放射性同位素标记法在胸肌组织的肌细胞和脂肪细胞中体外检测从头生成的FAs(n=3个细胞重复)。b.不同发育阶段肌肉组织中丙二酸-CoA的含量(n=28)。c.和d.肌肉组织和肝脏中丙二酸-CoA含量和FASNmRNA水平的比较(n=28)。e.和f.肌肉组织中肌细胞和脂肪细胞中丙二酸-CoA含量和FASNmRNA水平的比较。*,**,***。
4FASN通过DNL促进IMF沉积
为了验证FASN基因编码的FASN蛋白在DNL产生IMF中的功能,研究人员进行了更多的实验。不出所料,基因表达分析表明,GG纯合子中的FASN表达水平显著高于AA纯合子中的FASN表达水平(图4a),这与之前报道的胸部肌肉组织中FA含量与IMF/TG含量的相关性较高是一致的。研究人员还通过转录组和表型数据(包括IMF、TGs和FAs)对胸肌肌肉组织中表达的基因(n=14,259个)进行了WGCNA分析。通过差异分层聚类分析获得了基因模块,合并后共确定了22个模块。FASN基因出现在显著富集的ME淡黄色模块中,该模块与IMF、TG、C14:0、C14:1、C16:0、C16:1、C18:1n9c、C18:3n3和C20:1性状相关。研究结果表明,FASNmRNA水平与TG含量(r=0.526,P=0.044),C14:0(r=0.658,P=0.0076)之间存在显著的相关性。如图4b所示。此外,在ME浅黄色模块中还发现了521个与脂质代谢相关的代表性基因,这些基因被认为是FASN的共表达基因,主要参与FA代谢(ACLY和ACSS1)、TG脂肪分解(DGKA和LPL)和酯化(PLIN4)。此外,研究人员通过分析过表达FASN的鸡胸肌肌细胞,进一步研究了FASN对DNL的调控作用,发现随着FASN水平的升高,C14:0和TG的含量显著增加(图4c,4d和4e)。
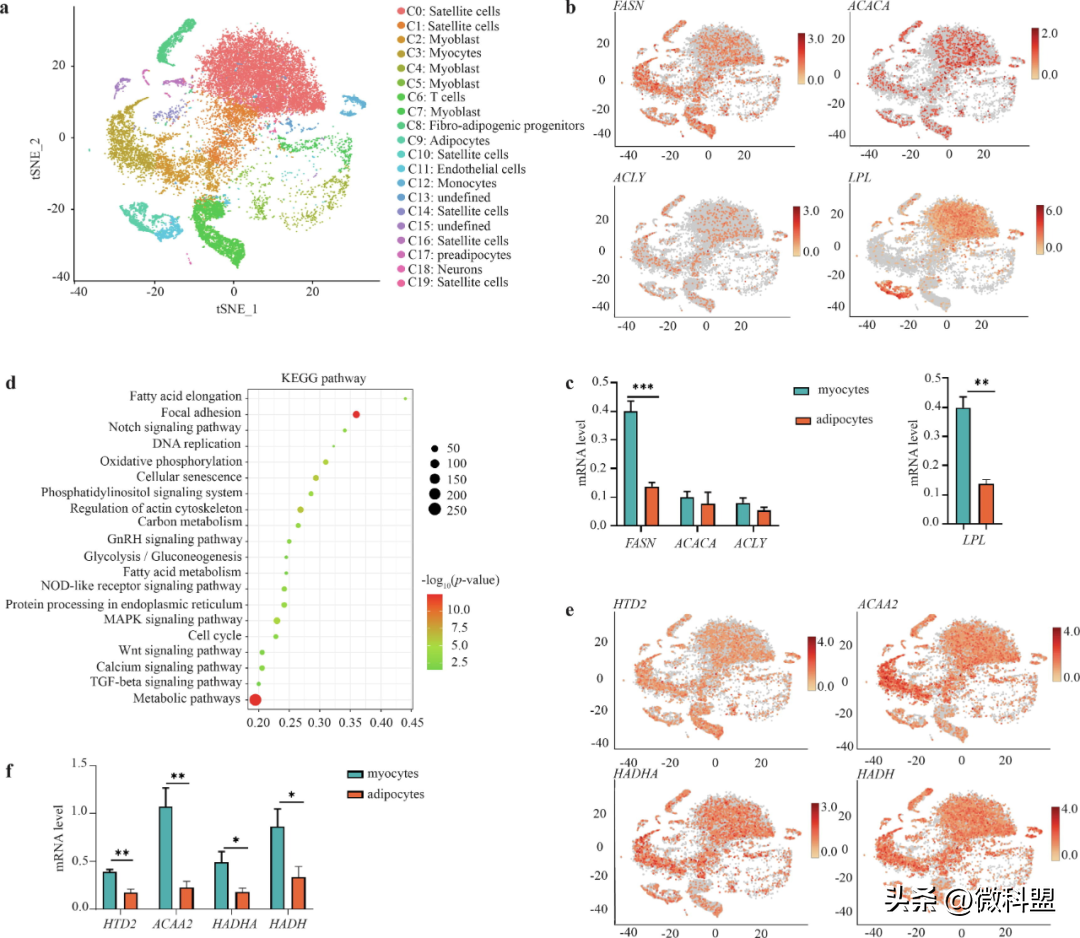
图4|AAA香蕉的基因组进化FASN通过DNL对IMF沉积的影响。a.携带rs315349829的GG或AA基因的纯合子(n=12)的胸肌组织中FASN的表达水平。及与FASN共表达的脂质代谢相关代表基因的表达水平与胸肌组织中FAs和TG含量的关系(n=16)。通过WGCNA检测胸肌组织中FASN的表达水平与FAs、TG含量(n=16)之间的关系,以及与FASN共表达的脂质代谢相关的代表性基因与胸肌组织中FAs、TG含量之间的关系。:0和TG含量随FASN表达水平增加的变化(n=3个细胞重复)。*,**,***。
5FASN在鸡肌肉组织的各类细胞中广泛表达
研究人员进行了单细胞RNA测序,以更准确地确定DNL的细胞学基础。63日龄时三个鸡胸肉样本的细胞活性见补充表16。单个样本过滤阈值为线粒体基因表达比例低于20%,细胞表达基因数为200-5000个。详细的统计结果,如每个细胞的基因中位数、独特分子标记(uniquemolecularidentifiers,UMI)、条形码中的Q30碱基和有效条形码,见补充表17和补充图10。通过参考其他物种数据库的标记基因,共系统地鉴定出20个细胞亚群,包括肌肉细胞群、脂肪细胞、单核细胞、T细胞、神经元等(图5a)。其中,与肌细胞和脂肪细胞相关的群体分别约占68.12%和10.37%。此外,研究人员还测定了一些代表性基因在不同细胞类型中的表达水平(图5b),发现参与DNL的基因(FASN、ACACA)在所有细胞类型中都有表达,而在肌肉细胞群中的信号更强。同样,参与脂肪分解的基因(如LPL)在所有细胞类型中都有表达。
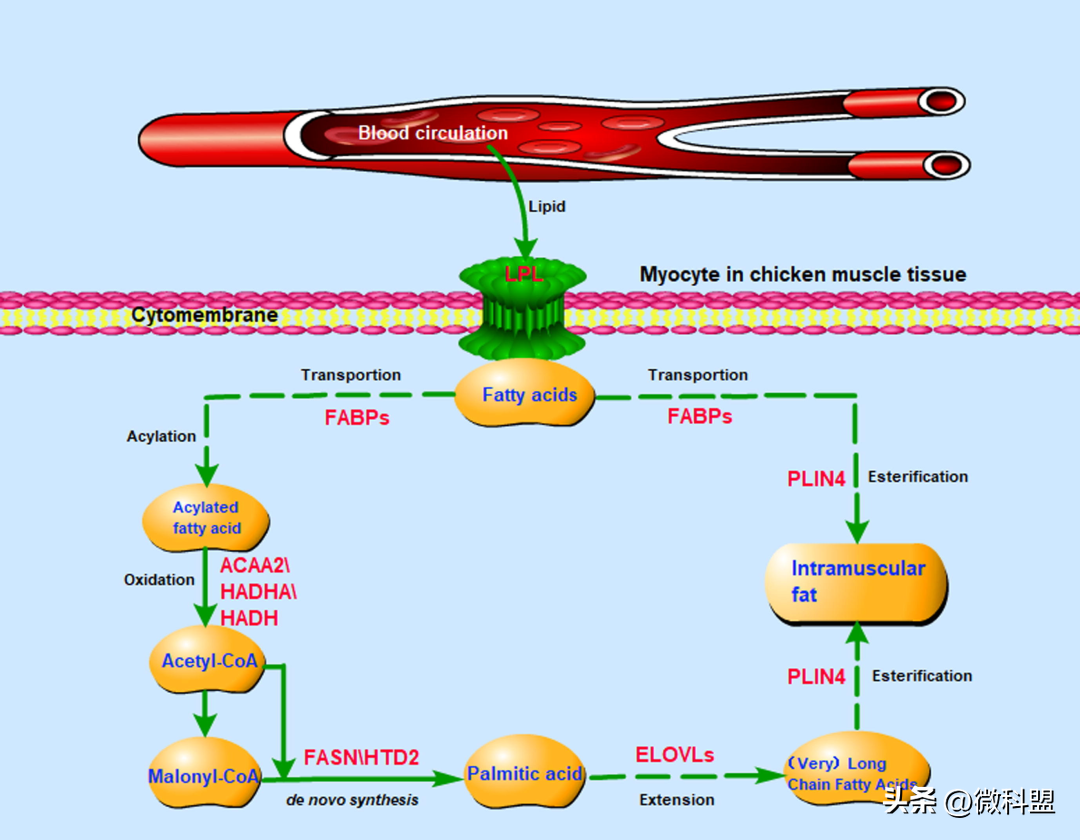
图5|FASN在肌肉组织DNL中的关键调控作用。a.通过单细胞测序鉴定的胸肌组织中的20个细胞亚群,主要包括肌肉细胞群和脂肪细胞,分别占68.12%和10.37%。b.和c.不同细胞类型中与DNL(FASN和ACACA)和脂肪分解(如LPL)相关的代表性基因的表达水平,以及与胸肌组织中的脂肪细胞相比,肌细胞中FASN的表达上调,LPL的表达下调(n=3)。d.基于3483个DEGs的KEGG通路富集分析对FA代谢通路的富集。e和f.除FASN外,HTD2、ACAA2、HADHA和HADH的表达水平在不同细胞类型中与FA代谢相关。
6FASN仍是调节肌肉组织中DNL的关键基因
针对上述检测到的胸肌组织中肌细胞和脂肪细胞中DNL水平的差异,研究人员还对差异表达基因(differentiallyexpressedgenes,DEGs,肌细胞对脂肪细胞)进行了比较分析,共鉴定出3483个DEGs(倍数变化1.5或0.67,P值0.05),其中包括1591个上调基因和1892个下调基因。值得注意的是,与脂肪细胞相比,肌细胞中FASN的表达水平显著上调,而LPL的表达水平显著下调(图5c)。此外,对3483个DEGs的KEGG通路富集分析显示,FA代谢是一个关键的显著富集通路(图5d),由FASN、HTD2、ACAA2、HADHA、HADH等编码的蛋白介导(图5e和5f)。在FA代谢途径中,FAs的从头合成是由FASN基因编码的酶催化的。在FA代谢途径中,FASN基因编码的酶以FA氧化产生的乙酰-CoA或丙二酰-CoA为底物催化FA的从头合成,并延伸为各种超长链FAs。
讨论
家禽是世界上产量和消费量最大的肉类,其消费量取决于其美味、鲜嫩和多汁的品质。已经证实,IMF含量高是影响肉质风味、质地和其他理想特性的重要因素,因此提高IMF含量是改善肉质的有效方法。然而,鸡肉中IMF产量的遗传基础仍不清楚。因此,为了更好地了解IMF产生的遗传基础并阐明鸡IMF沉积的机制,研究人员利用IMF含量高的表型实验模型进行了一项系统研究。这项研究不仅揭示了编码负责将葡萄糖转化为FAs的多酶蛋白的FASN基因是促进鸡体内IMF沉积的主要基因,而且还揭示了SNPrs315349829可能是导致肌肉组织中C14:0、C16:0和TG含量增加的因果突变,这将有助于指导优质鸡肉的育种策略。
对鸡胸肌组织中IMF含量的GWAS分析表明,IMF是一个受多个基因和基因网络调控的复杂性状,其中涉及与FA代谢相关的基因,这一点得到了RNA测序和单细胞测序数据的支持。这些发现也与研究人员之前报道的结果一致。随后,研究人员利用多组学组合方法研究了胸肌组织中的FA代谢,重点阐明了鸡IMF含量这一性状的遗传基础,该方法在系统筛选与数量性状相关的关键功能基因的遗传变异方面具有不可否认的优势。研究人员在FASN的上游区域发现了SNPrs315349829,它可能通过上调FASN在鸡胸肌组织中的表达水平而导致C14:0和C16:0含量的增加。这些结果表明,与对照组相比,SNPrs315349829和FASN基因可能是导致IMF选择的鸡群体胸肌中IMF含量增加的原因。鉴于C14:0和C16:0分别是FA从头生物合成的中间产物和最终产物,而FASN编码驱动DNL的关键酶,研究人员假设DNL可能是促进鸡体内IMF沉积的重要因素,正如研究人员先前的研究所建议的那样。然而,目前流行的理论认为,家禽的DNL主要发生在肝脏中,而肌肉组织和脂肪组织中几乎没有。因此,为了证实研究人员关于DNL对IMF沉积的贡献的假设,研究人员进行了更多的体内和体外同位素追踪(使用金标记法)实验,检测丙二酰-CoA(合成脂肪酸的不可逆第一中间体)的存在以及FA在鸡IMF沉积过程中的从头合成。
一般认为,大多数IMF储存在脂肪细胞中,而不是在肌细胞中,除非在病理条件下发生异位沉积。令人惊讶的是,尽管有报道称鸡肌肉组织中的IMF含量很低(仅为2%-4%),但在肌细胞中,新合成的同位素标记的FAs的比例被测定为总FAs的35%-40%。因此,研究人员的研究结果表明,肌肉组织的肌细胞和脂肪细胞中都存在DNL,但主要发生在肌细胞,这符合肌细胞的生理特点,肌细胞的能量需求主要通过细胞摄取的FAs氧化降解(分解代谢)来满足,多余的能量可以储存在附近。研究人员利用单细胞测序技术获得的数据显示,FASN基因在肌肉细胞群中的表达量最高,这再次证实了鸡肌肉组织中存在DNL。正如预期的那样,研究结果表明,根据肌细胞和脂肪细胞产生DNLFA的差异,FA代谢途径得到了丰富,FASN以乙酰-CoA或丙二酰-CoA为底物促进了DNL的产生。此外,其他已鉴定的DEGs(ACAA2、HAD-HA和HADH)编码的其它酶促进肉豆蔻酸的非从头合成,肉豆蔻酸降解为乙酰-CoA,为DNL提供起始底物。换句话说,肌肉组织利用细胞摄取的FAs氧化产生的能量,并利用多余的能量通过DNL再生FAs。基于上述结果,研究人员提出了鸡肌肉组织中IMF沉积的潜在调控机制(图6)。
图6|鸡肌肉组织中IMF沉积的潜在调控机制。肌肉组织使用通过FA细胞摄取获得的FA氧化产生的能量(ACAA2、HADHA和HADH),多余的能量则用于通过DNL(FASN和HTD2)再生FA。